
El núcleo es el lugar de la célula que contiene el material genético y dirige toda la actividad celular.
En el núcleo interfásico —estructura que presenta el núcleo cuando no se divide—, podemos observar una envoltura nuclear, que encierra al nucleoplasma, en el que se encuentran el nucleolo y la cromatina.
a) Envoltura nuclear
Procede del retículo endoplasmático y rodea al núcleo separándolo del resto del citoplasma. Es una envoltura doble formada por dos membranas: la membrana nuclear externa —que puede llevar unidos ribosomas en la cara que da al citosol— y la membrana nuclear interna. Entre ambas queda un pequeño espacio denominado espacio perinuclear, que se continúa con el espacio del retículo.
La membrana nuclear interna lleva asociada a su cara nucleoplásmica una red de filamentos proteicos que forman la lámina fibrosa o corteza nuclear. Su función es participar en la formación de la envoltura nuclear después de la mitosis y en la organización de la cromatina.
La envoltura nuclear no es continua, pues existen puntos en los que las dos membranas se unen creando unos orificios denominados poros nucleares.
Los poros nucleartes, con un diámetro de entre 50-100 nm, están formados por una estructura compleja llamada complejo del poro, constituida por ocho masas proteicas que se disponen en forma de octógono y forman un anillo o cilindro hueco que reviste internamente el poro. Asociado al anillo se encuentra un material denso, el diafragma, que disminuye la luz del poro hasta 10 nm.

Los poros nucleares regulan el intercambio de moléculas entre el núcleo y el hialoplasma.
b) Nucleoplasma
También se le denomina matriz nuclear. Es el medio interno del núcleo —similar al hialoplasma—, formado por una disolución coloidal compuesta por agua, iones, numerosas proteínas —histonas que intervienen en el empaquetamiento del ADN, enzimas que intervienen en la replicación y transcripción del ADN—, nucleótidos necesarios para la síntesis de los ácidos nucleicos, etcétera.
c) Nucleolo
Es una estructura esférica, no rodeada de membrana, densa y con un contorno irregular. Su función es fabricar los distintos tipos de ARN ribosómico que forman parte de las subunidades de los ribosomas.
Se encuentra formado por ARN, ADN y proteínas, y en él se distinguen, al microscopio electrónico, tres zonas:
Zona fibrilar: zona más interna, formada por bucles de ADN que llevan información para sintetizar ARNn (nucleolar); a estos fragmentos se les denomina organizadores nucleolares. Estos fragmentos pueden pertenecer a uno o a varios cromosomas diferentes, que se denominan cromosomas organizadores del nucleolo.
Componente fibrilar denso: lugar del nucleolo donde el ADN organizador nucleolar de cada cromosoma empieza a transcribirse. Una vez sintetizadas las copias de ARN nucleolar 45s, se unen a proteínas formando fibrillas, que serán parte de los ribosomas.
Zona granular: zona más periférica, que contiene las subunidades ribosómicas en proceso de maduración. Estas subunidades saldrán al citoplasma a través de los poros nucleares; allí terminan de madurar y se unen a los ARN mensajeros, formando polirribosomas.
d) Cromatina
Se denomina así al material genético de la célula eucariota durante la interfase.
Con el microscopio óptico aparece como masas densas que se distribuyen por el núcleo.
Con el microscopio electrónico se observa que tiene estructura fibrilar formada por filamentos que se entrecruzan entre sí, formando un retículo que está inmerso en el nucleoplasma; al comenzar la división celular estos filamentos se condensan dan lugar a los cromosomas.
La cromatina están formada por ADN bicatenario lineal que está asociado a proteínas histonas, que son proteínas básicas —ricas en aminoácidos básicos: arginina y lisina— de bajo peso molecular. Además, hay otras proteínas no histónicas, en su mayoría enzimas que intervienen en la transcripción y replicación del ADN.
Las fibras de cromatina presentan distintos niveles de organización que facilitan su empaquetamiento: nucleosoma, collar de perlas, fibras de 30nm (300A).

Durante la interfase pueden diferenciarse distintos tipos de cromatina:
Eucromatina: zonas donde la cromatina está poco condensada. Está formada por los fragmentos de ADN correspondientes a los genes activos (transcriben ARNm) así como los fragmentos de ADN que llevan información para la transcripción del ARNt y ARNr.
Heterocromatina: zonas donde la cromatina está muy condensada y por lo tanto se tiñe fuertemente, representa el 90%. Se corresponde con las zonas en las que el ADN no se transcribe y permanece funcionalmente inactivo durante la interfase.

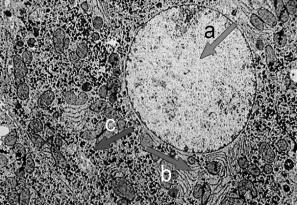
Fig. 10.7 - Microfotografía electrónica del núcleo de un hepatocito. a. eucromatina; b. RER

Fig. 10.8 - Microfotografía electrónica del núcleo de un linfocito. a. eucromatina; b. heterocromatina, c. mitocondria
CROMOSOMAS Y CROMATINA
Cuando el cromosoma en interfase se esparce artificialmente sobre agua, tiene la apariencia de un collar de perlas. Las perlas son los nucleosomas, las unidades de enrollamiento de la cromatina. (Fig. 10.10b)
Los nucleosomas están formados por un centro o "core" de histonas. Dicho centro posee dos copias de cada una de las siguientes histonas: H2A; H2B; H3 y H4
Alrededor del centro de histonas, 146 pares de bases del ADN se enrollan en dos vueltas. La unión de las histonas al ADN no depende de una secuencia particular de nucleótidos, sino de la secuencia de aminoácidos de la histona. Las histonas son unas de las moléculas más conservadas durante el transcurso de la evolución. La histona H4 en el ternero difiere de la H4 de la planta de poroto en sólo 2 residuos de aminoácidos de una cadena de 102. Alrededor de 60 pares de bases de ADN unen un nucleosoma con el próximo. Cada región de unión es el ADN espaciador. La quinta histona, la H1, conecta a los nucleosomas y actúa como una banda de goma, manteniéndolos juntos dentro de una misma cuerda enrollada. Esta estructura se conoce como fibra de 10nm, siendo el primer grado del empaquetamiento de la cromatina. (Fig. 10.9)
Los nucleosomas se organizan, a su vez, en fibras de 30nm (solenoide), girando a manera de resorte alrededor de un eje virtual. Esta estructura es mantenida por la interacción de las H1 de nucleosomas cercanos. (Fig. 10.10c)
En el siguiente nivel de empaquetamiento, las fibras de 30 nm se organizan en una serie de bucles o asas superenrolladas (Fig. 10.10d; Fig. 10.11). Estos bucles se estabilizan gracias a la interacción con las proteínas de la matriz nuclear o andamiaje nuclear (“scaffold”).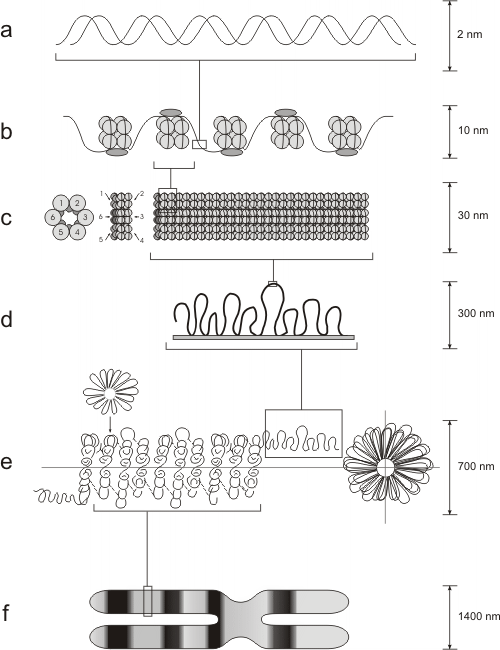
Fig. 10.7 - Microfotografía electrónica del núcleo de un hepatocito. a. eucromatina; b. RER
Fig. 10.8 - Microfotografía electrónica del núcleo de un linfocito. a. eucromatina; b. heterocromatina, c. mitocondria
CROMOSOMAS Y CROMATINA
El núcleo contiene los cromosomas de la célula. Cada cromosoma consiste en una molécula única de ADN con una cantidad equivalente de proteínas. Colectivamente, el ADN con sus proteínas asociadas se denomina cromatina. La mayor parte de las proteínas de la cromatina consisten en copias múltiples de cinco clases dehistonas.
Estas proteínas básicas son ricas en residuos de arginina y lisina cargados positivamente. Por esta razón se unen estrechamente con los grupos fosfatos (cargados negativamente) del ADN.
La cromatina también contiene pequeñas cantidades de una amplia variedad de proteínas no histónicas y RNP. La mayoría de ellas son factores de transcripción (por ej., el receptor esteroide), siendo su asociación con el ADN pasajera. Estos factores regulan que parte del ADN será transcripta en ARN.
NIVELES DE ORGANIZACIÓN DE LA CROMATINA
La observación a través del microscopio óptico de un núcleo interfásico, nos permite distinguir dos tipos de cromatina. La eucromatina o cromatina laxa, de localización central, y la heterocromatina o cromatina densa, en la periferia del núcleo (Fig.10.7). La heterocromatina representa aproximadamente el 10% del total de cromatina y es considerada transcripcionalmente inactiva (Fig. 10.8).
La eucromatina se encontraría al menos en dos estados, la eucromatina accesible, que representa alrededor del 10%, donde se encuentran los genes que se están transcribiendo y la eucromatina poco accesible, más condensada (pero menos que la heterocromatina), donde están los genes que la célula no está transcribiendo.Cuando el cromosoma en interfase se esparce artificialmente sobre agua, tiene la apariencia de un collar de perlas. Las perlas son los nucleosomas, las unidades de enrollamiento de la cromatina. (Fig. 10.10b)
Los nucleosomas están formados por un centro o "core" de histonas. Dicho centro posee dos copias de cada una de las siguientes histonas: H2A; H2B; H3 y H4
Alrededor del centro de histonas, 146 pares de bases del ADN se enrollan en dos vueltas. La unión de las histonas al ADN no depende de una secuencia particular de nucleótidos, sino de la secuencia de aminoácidos de la histona. Las histonas son unas de las moléculas más conservadas durante el transcurso de la evolución. La histona H4 en el ternero difiere de la H4 de la planta de poroto en sólo 2 residuos de aminoácidos de una cadena de 102. Alrededor de 60 pares de bases de ADN unen un nucleosoma con el próximo. Cada región de unión es el ADN espaciador. La quinta histona, la H1, conecta a los nucleosomas y actúa como una banda de goma, manteniéndolos juntos dentro de una misma cuerda enrollada. Esta estructura se conoce como fibra de 10nm, siendo el primer grado del empaquetamiento de la cromatina. (Fig. 10.9)
Los nucleosomas se organizan, a su vez, en fibras de 30nm (solenoide), girando a manera de resorte alrededor de un eje virtual. Esta estructura es mantenida por la interacción de las H1 de nucleosomas cercanos. (Fig. 10.10c)
En el siguiente nivel de empaquetamiento, las fibras de 30 nm se organizan en una serie de bucles o asas superenrolladas (Fig. 10.10d; Fig. 10.11). Estos bucles se estabilizan gracias a la interacción con las proteínas de la matriz nuclear o andamiaje nuclear (“scaffold”).
Los cromosomas se diferencian por la ubicación del centrómero
Durante la mayor parte de la vida de la célula, los cromosomas son demasiado largos y tenues para ser vistos bajo un microscopio.

A primera vista, los cromosomas duplicados se mantienen juntos por el centrómero. Mientras están juntos, es común llamar a cada parte del cromosoma duplicado, cromátida hermana.
El cinetocoro es una estructura proteica discoidal que forma parte del centrómero y ayuda a separar las cromátidas hermanas. Es el sitio de unión con los microtúbulos del huso, que contienen los motores de dineína que tiran a los cromosomas en la anafase. Además proveen una plataforma para ensamblar y movilizar las proteínas que construyen el huso.
La posición del centrómero, determina el largo de los brazos del cromosoma; en base a esto se puede clasificar a los cromosomas en: (Fig. 10.15)
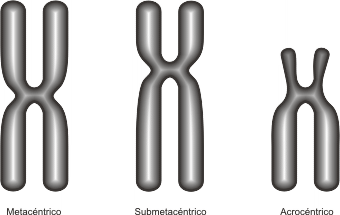
Fig. 10.15- Tipos de cromosomas
· Metacéntricos: el centrómero en posición central determina brazos de igual longitud
· Submetacéntricos: un par de brazos es más corto que el otro, pues el centrómero se encuentra alejado del centro.
· Acrocéntricos: el centrómero se halla próximo a uno de los extremos, por lo tanto uno de los brazos es casi inexistente.
Todas las especies tienen un número característico de pares de cromosomas homólogos llamado número diploide (2n). El número diploide del hombre es 46.
El cariotipo es una representación gráfica o fotográfica de los cromosomas presentes en el núcleo de una sola célula somática de un individuo. Cada miembro del par de cromosomas homólogos proviene de cada uno de los padres del individuo cuyas células examinamos.
El cariotipo de la mujer contiene 23 pares de cromosomas homólogos, 22 pares son autosomas y el par restante, cromosomas sexuales, ambos " X".
El cariotipo del hombre contiene los mismos 22 pares de autosomas y 1 par de cromosomas sexuales, un cromosoma sexual "X" y un cromosoma sexual "Y" (un gen en el cromosoma Y designado SRY es el que pone en marcha el desarrollo de un varón, por lo tanto determina el sexo).
El análisis del cariotipo involucra la comparación de cromosomas por su longitud, la ubicación de los centrómeros y la ubicación y los tamaños de las bandas G.
Durante la mitosis, los 23 pares de cromosomas humanos se condensan y son visibles con un microscopio óptico.
Preparación de un Cariotipo: La preparación de un cariotipo normalmente involucra bloquear las células (glóbulos blancos) durante la mitosis con colchicina y marcar los cromosomas condensados con tinción Giemsa. La tinción marca las regiones de los cromosomas que son ricos en pares de nucleótidos entre A -T produciendo una banda oscura, la banda G. Luego de la tinción, los cromosomas se fotografían, se recortan y se ordenan de acuerdo a su longitud. Los de igual tamaño se aparean según la ubicación de su centrómero.
Una error común es suponer que cada banda representa un sólo gen. En realidad las bandas más pequeñas contienen más de un millón de pares de nucleótidos y potencialmente cientos de genes. Por ejemplo, el tamaño de una banda pequeña es igual a toda la información genética de una bacteria.
El análisis del cariotipo es una de muchas técnicas que nos permiten investigar las miles de enfermedades genéticas que se pueden encontrar en los seres humanos.
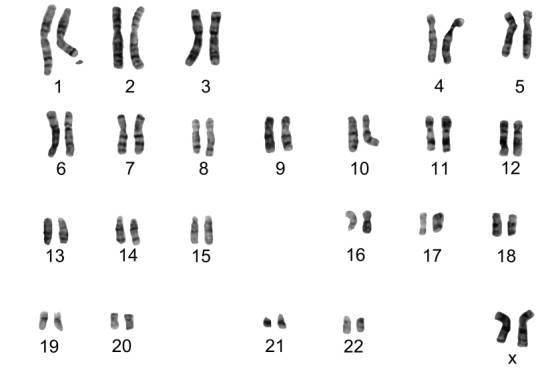
Fig. 10.17 - Cariotipo femenino normal: Los autosomas se ordenan en grupos por tamaño y posición del centrómero.
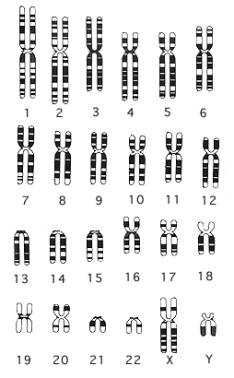 Cariotipo masculino normal.
Cariotipo masculino normal.
No hay comentarios:
Publicar un comentario